LifeTein successfully synthesized Aureocin A53, a highly cationic 51-residue peptide containing ten lysine and five tryptophan residues, Formyl-MSWLNFLKYIAKYGKKAVSAAWKYKGKVLEWLNVGPTLEWVWQKLKKIAGL, using solid-phase peptide synthesis approach. A53 is a Class II bacteriocins. It was originally isolated from Staphylococcus aureus A53 and is active against methicillin-resistant Staphylococcus aureus (MRSA) and vancomycin-resistant Enterococcus. The mode of action was proposed as insertion into lipid bilayer and consequent membrane leakage. The solid-phase peptide synthesis provide an inexpensive alternative to isolation from bacteria culture or recombinant expression.
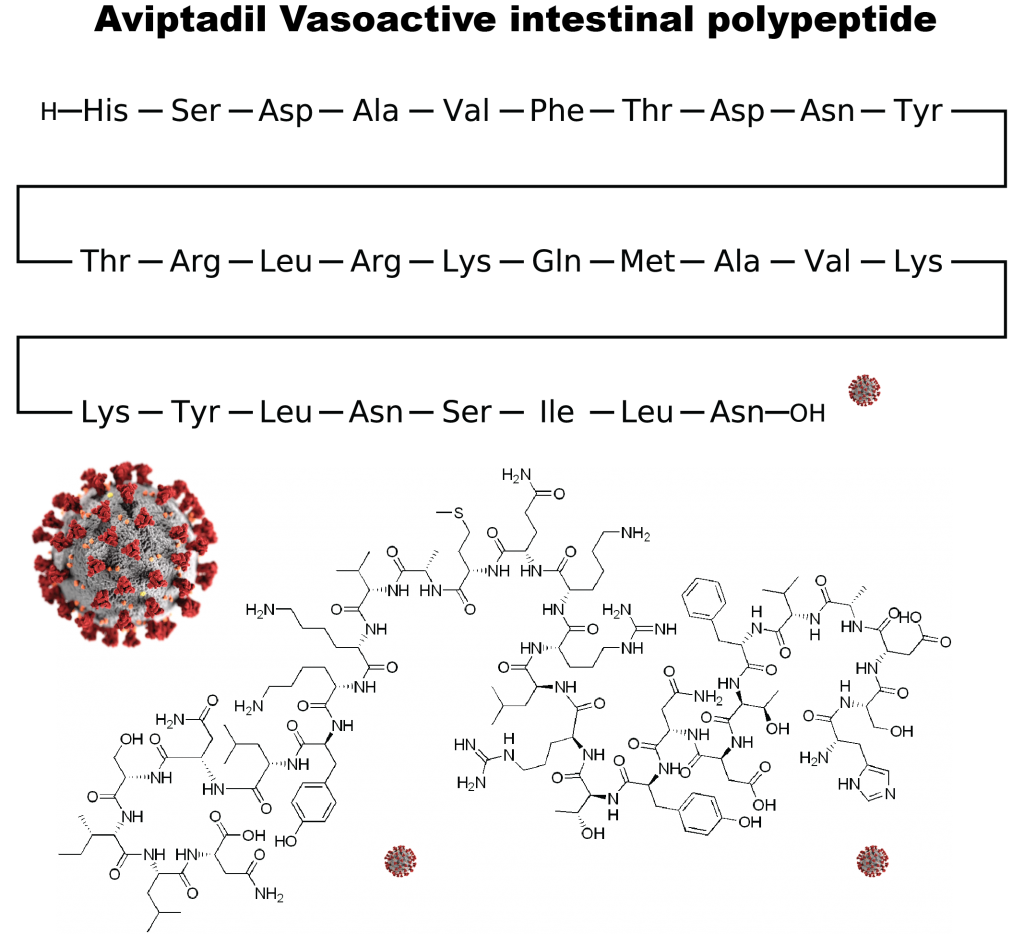
It was found that RLF-100 (Aviptadil) is associated with rapid respiratory failure recovery among COVID-19 Patients. The clinical findings may be based on evidence that VIP inhibits the replication of the SARS-CoV-2 virus in human lung cells and immune cells (monocytes). No other antiviral agent has demonstrated rapid recovery from viral infection and demonstrated laboratory inhibition of viral replication. It is a patented formulation of aviptadil (synthetic human Vasoactive Intestinal Polypeptide, VIP), which has been granted FDA fast track designation, FDA emergency use IND authorization, and an expanded access protocol. Aviptadil is an injectable formulation of the vasoactive intestinal polypeptide (VIP) in combination with the adrenergic drug phentolamine.
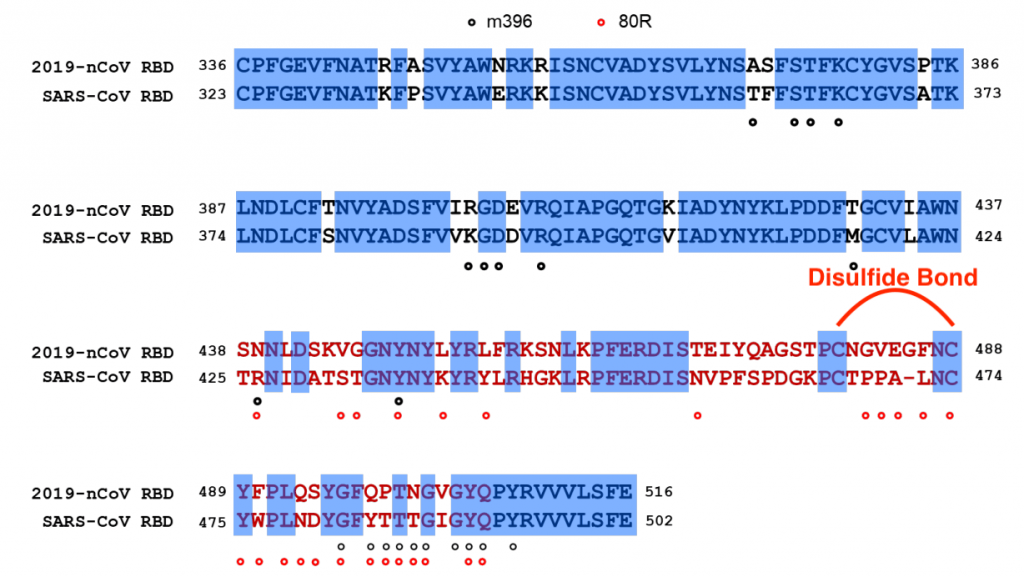
Coronavirus receptor binding domain. The key receptor binding domain (residues 319-541) is highlighted in yellow. Variable amino acid residues between SARS-CoV-2 and SARS-CoV are highlighted in cyan. Tyr 489, Asn 487, Gln 493, Tyr 505 are important for ACE2 binding.
LifeTein can help in your research with custom peptide synthesis of the following specific proteins: SARS-CoV-2 Receptor Binding Domains, SARS-CoV-2 Nucleocapsid Fragments, T-cell and B-cell Epitopes of SARS-CoV-2, Fusion Inhibitors Targeting HR1 Domain of the SARS-CoV-2 Spike Proteins, Inhibitors of SARS-CoV-2 Mpro/3CLpro/C30 Endopeptidase, ACE2 Inhibitors and Substrates, and AT2 Receptor Agonists and Antagonists.
Pool of 22 peptides derived from a peptide design (15mers with 5 aa overlap) through the receptor binding domain of S1 protein.
Direct antimicrobial mechanisms of cationic host defense peptides
The cationic host defense peptides (CHDP), also known as antimicrobial peptides, could be used to kill enveloped viruses such as the 2019 Novel Coronavirus SARS-CoV-2. The peptides have the potential to destabilize the viral envelope on contact, damaging the virions and inhibiting infectivity. The specific antiviral peptide may bind to cellular receptors involved in viral infection or peptide-mediated aggregation of viral particles. The antiviral peptides could create an ‘antiviral shield’ at mucosal surfaces and prevent replication and spread of the Coronavirus if upregulated after the initial infection.
During pandemics, where there is insufficient time to produce vaccines (such as the outbreak of respiratory illness Covid-19 first detected in Wuhan, China), the cationic host defense peptides could be the first-line antiviral treatments.
Some of the antimicrobial peptides are the human cathelicidin LL-37 and β-defensins. Cathelicidins are immunomodulatory antimicrobials with an important role in the regulation of the inflammatory response. The only human cathelicidin, LL-37, is the most well-studied peptide in this family. LL-37 is an α-helical peptide. While defensins have a common β-sheet core stabilized with three disulfide bridges between six conserved cysteine residues.
Direct antimicrobial mechanisms of cationic host defense peptides can be mediated by membrane translocation of the peptides followed by binding to intracellular targets such as nucleic acids and/or proteins to kill bacteria. Proline-rich antimicrobial peptides use inner membrane transporters as Trojan horses to gain entry and bind to intracellular targets such as nucleic acids or nascent proteins. And subsequently affect cell processes such as replication, transcription, translation, protein folding, and cell wall synthesis.
At this stage, only a few peptide-derived treatments have made it to market such as PAC-113, a histatin analog, and dalbavancin, a semisynthetic lipoglycopeptide.
Despite the limited understanding of structure-function relationships, the potential of peptide-based therapies remains a promising new clinical direction for the Coronavirus.
The extensive structural analyses have revealed that interactions between SARS-CoV spike protein receptor-binding domain (RBD) and its host receptor angiotensin-converting enzyme 2 (ACE2), which regulate both the cross-species and human-to-human transmissions of SARS-CoV.
Studies showed that the sequence of 2019-nCoV coronavirus RBD, including its receptor -binding motif (RBM) that directly contacts ACE2 and uses ACE2 as its receptor with much higher affinity (10-20 times higher!) than SARS.
Several critical residues in 2019-nCoV RBM may provide favorable interactions with human ACE2 such as Gln493 and Asn501.
A total of nine cysteine residues are found in the RBD, six of which forming three pairs of disulfide bonds. Among these three pairs, two are in the core (Cys336-Cys361 and Cys379-Cys432) to help stabilize the β sheet structure while the remaining one (Cys480-Cys488) connects loops in the distal end of the RBM.
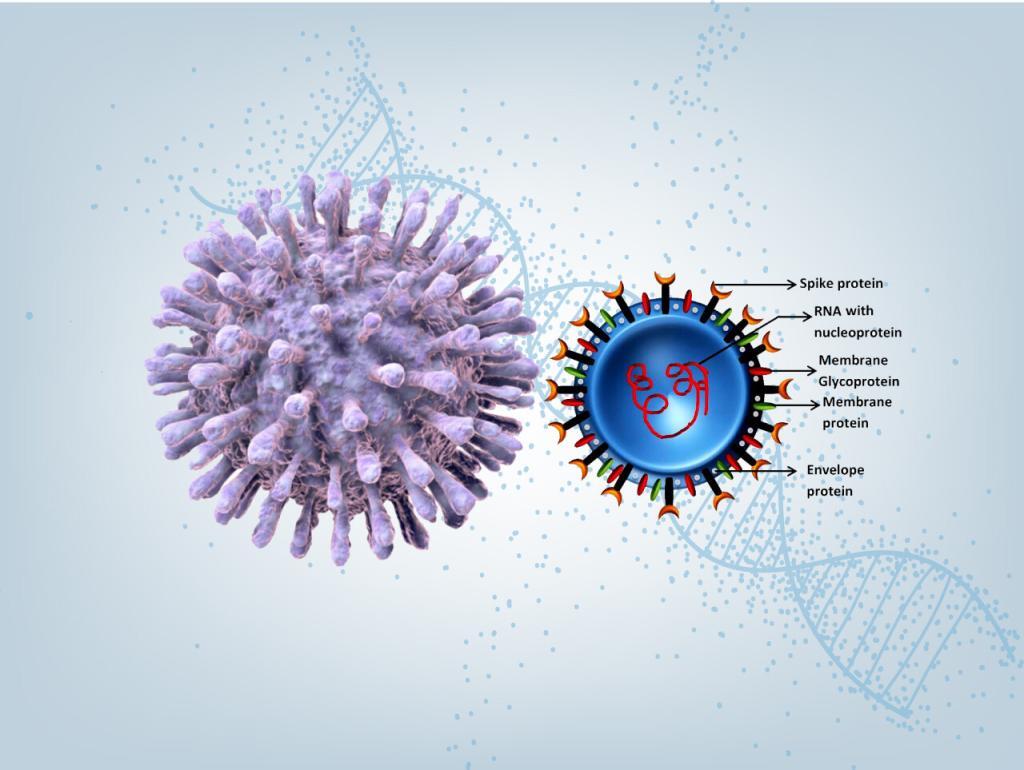
2019 Novel Coronavirus SARS-CoV-2 is a virus identified as the cause of an outbreak of respiratory illness Covid-19 first detected in Wuhan, China.
To help expedite Covid-19 research, LifeTein synthesized a 69 amino acid spike glycoprotein with one disulfide bond in 6 days. This effort is a partnership with a biotech company for drug development.
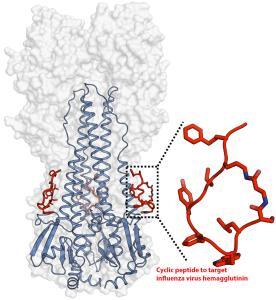
Cyclic peptides as broad-spectrum antiviral agents
Antiviral drugs and vaccines are the most powerful tools to combat viral diseases. Most drugs and vaccines only target a single virus. However, the broad-spectrum antivirals can be used for rapid management of new or drug-resistant viral strains. Cyclized peptides and peptide analogs are excellent examples of broad-spectrum antivirals.
An artificial peptide molecule was found to neutralize a broad range of group 1 influenza A viruses, including H5N1. The peptide design was based on complementarity determining region (CDR) loops have been reported for other viral targets. The optimized peptides bind to the highly conserved stem epitope and block the low pH-induced conformational rearrangements associated with membrane fusion.
These peptidic compounds and their advantageous biological properties should accelerate development of novel small molecule and peptide-based therapeutics against influenza virus.
The linear peptide is Suc-SQLRSLEYFEWLSQ-NH2. Three cyclization strategies were used: head to tail, side chain to side chain and side chain to tail. An ornithine (Orn) side chain was fused with the carboxyl terminus of β-alanine for lactam formation.
Check here for more details: Potent peptidic fusion inhibitors of influenza virus, Science 28 Sep 2017, DOI: 10.1126/science.aan0516
Lately, more broad-spectrum antiviral agents were found to target viruses. It was found that 55 compounds can target eight different RNA and DNA viruses. Dalbavancin is a novel lipo-glycopeptide antibiotic. The lipoglycopeptide disrupts bacterial cell wall formation by binding to
the terminal d-alanyl-d-alanine peptidoglycan sequence in Gram-positive bacteria in a linear, concentration-dependent manner. The dalbavancin has effects on echovirus 1, ezetimibe against HIV1 and Zika virus.
More details: https://www.ncbi.nlm.nih.gov/pubmed/29698664
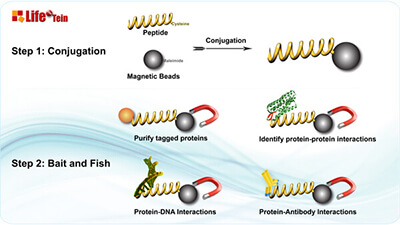
Protein Purification Using Magnetic Beads: Top Tips for Success
Magnetic bead-based protein purification offers a powerful solution for various applications like high-throughput microscale purification, pull-down/CoIP experiments, and protein-protein or protein-DNA interaction studies. Here’s why magnetic beads are the top choice: they can be coated with specific affinity ligands for antigens, antibodies, proteins, or nucleic acids. Moreover, magnetic beads are non-porous and have a defined diameter, eliminating hidden surfaces where molecules can stick, leading to reduced background, simplified purification, and streamlined washing steps. Compared to traditional bead separation methods involving agarose, sepharose, or silica beads, magnetic bead separation stands out as the quickest, cleanest, and most efficient technique.
If you’re new to working with magnetic beads, here are some essential tips to ensure success:
Thorough Resuspension: Ensure uniformity across aliquots by thoroughly resuspending your magnetic beads. These nano-superparamagnetic beads are covalently coated with highly functional groups, providing increased binding capacity and better dispersion. Since magnetic beads are composed of iron oxide and can settle over time, it’s crucial to vortex and resuspend them thoroughly before use to redisperse the beads.
Enhanced Washing: Minimize non-specific binding by increasing the number of washing steps. Whether you’re using ethanol or the recommended wash buffer, make sure to use an adequate volume of wash solution to cover the bead pellet.
Understanding Functional Groups: Different beads are covalently coated with various functional groups like maleimide, primary amine, NHS, carboxylic acid, purified streptavidin, protein A, reduced glutathione, nickel-charged nitrilotriacetic acid, or groups for DNA/RNA purification. These coatings, along with buffer conditions, affect bead properties. Understanding these specifics is essential for proper bead handling.
Efficient Bead Capture: Magnetic beads typically form a pellet attracted to the magnet within a minute. Extend the attraction time to ensure efficient bead capture.
Gentle Supernatant Removal: When removing the wash solution or supernatant, angle the pipette tip to avoid disturbing the magnetic bead pellet. Ensure that the tip doesn’t come into contact with the pellet.
By following these tips, you can make the most of magnetic bead-based protein purification, improving the efficiency and reliability of your experiments.
Giantin, a novel conserved Golgi membrane protein, is a disulfide-linked homodimer. It was found that BFA-induced Golgi disorganization is associated with the monomerization of giantin.
The pull-down experiment was performed. The control peptide biotin-GHGTGSTGSGSMLRTLLRRRL synthesized by LifeTein was incubated with lysate and Dynabeads, as well as the lysate incubated with Dynabeads only served as a control. Dynabeads carrying MGAT1 peptide were able to pull-down giantin from the lysate of HeLa cells, however, giantin was not detected in the pull-down fraction from the lysate exposed to the Dynabeads or in combination with control peptide. It is logical to hypothesize that the MGAT1 binding domain of giantin lies within its N-terminal non-coiled-coil area.
The Dynabeads function similarly to LifeTein magnetic beads: https://www.lifetein.com/peptide-product/amineactivated-peptide-conjugation-magnetic-beads-p-3647.html
polycyclic CPKILKKCRRDSDCPGACICRGNGYCGSGSDGGV (Huang et al. 2015)
MFK
MFKLRAKIKVRLRAKIKL (Samuels et al. 2017)
Mgpe9
CRRLRHLRHHYRRRWHRFRC (Vij et al. 2016a)
MitP
INLKKLAKL(Aib)KKIL (Howl et al. 2018)
m(KLA)-iRGD
klaklakklakla-K-GG-iRGD (Qifan et al. 2016)
MMGP1
MLWSASMRIFASAFSTRGLGTRMLMYCSLPSRCWRK (Pushpanathan et al. 2013)
MPER fragment
ELDKWASLWNWFDITNWLWYIK (Song et al. 2009)
MPG
GALFLGFLGAAGSTMGA cysteamide (Morris et al. 1997)
MPG
GALFLGFLGAAGSTMGASQPKKKRKV cycteamide (Deshayes et al. 2005)
MPG-8
AFLGWLGAWGTMGWSPKKKRK (Crombez et al. 2009b)
mRVG
YTIWMPENPRPGTPCDIFTKSRGKRASNGGGRRRRRRRRR (Villa-Cedillo et al. 2017)
MT23
LPKQKRRQRRRM (Zhou et al. 2017)
mtCPP1
r-Dmt-OF (Cerrato et al. 2015)
MTM
AAVALLPAVLLALLAP (Fletcher et al. 2010)
MTD84
AVALVAVVAVA (Lim et al. 2014)
MTP
MLSLRQSIRFFK (Chuah et al. 2015a, b)
MTS
KGEGAAVLLPVLLAAPG (Zhao et al. 2001)
MTS1
AAVLLPVLLAAP (Rojas et al. 1998)
Mut3DPT-C9h
VKKKKIKAEIKIYVETLDDIFEQWAHSEDL (de la Torre et al. 2017)
Myr-ApoE
Myr-LRKLRKRLLR (Tajik-Ahmadabad et al. 2017)
New modalities
Polycyclic, hairpin, stapled peptides for delivery (Valeur et al. 2017, Waldmann et al. 2017)
NF1
Stearyl-AGY(PO3)LLGKTNLKALAALAKKIL (Arukuusk et al. 2013)
NF51
δ-(Stearyl-AGYLLG)OINLKALAALAKKIL (Arukuusk et al. 2013)
NF55
δ-(Stearyl-AGYLLG)OINLKALAALAKAIL (Freimann et al. 2016)
NLS
PKKKRKV (Yoneda et al. 1992).
NLS-StAx-h
stapled RRWPRXILDXHVRRVWR (Dietrich et al. 2017)
NoLS
KKRTLRKNDRKKRC (Yao et al. 2015)
Novicidin
KNLRRIIRKGIHIIKKYF (Milosavljevic et al. 2016)
NPFSD
VLTNENPFSDP (Gong et al. 2016)
NYAD-1
stapled ITFEDLLDYYGP (Zhang et al. 2008)
Oct4-PTD
DVVRVWFCNRRQKGKR (Adachi et al. 2017)
P007
Ac-(RAhxR)4-Ahx-βAla (Greer et al. 2014)
P1
LRRWSLG (Peng et al. 2017b)
P2
WKRTLRRL (Peng et al. 2017b)
P3
YGRKKRRQR (Tan et al. 2006)
P7
RRMKWKK (Watson et al. 2017)
P11
YGRKKRRQRRR (Zhao et al. 2011)
P11
HSDVHK (Bang et al. 2011)
P11LRR
P11LRR (Li et al. 2010)
P14LRR
(PLPRPR)4 (Brezden et al. 2016)
p18
LSTAADMQGVVTDGMASG (Taylor et al. 2009)
P21
KRKKKGKGLGKKRDPCLRKYK (Dixon et al. 2016)
P28
LSTAADMQGVVTDGMASGLDKDYLKPDD, Leu50-Asp77 of azurin (Yamada et al. 2016)
p28
FLHSGTAVTCTYPALTPQWEGSDCTHRL (Signorelli et al. 2017)
p53 peptide MO6
Stapled TSF*EYWYLL* (Chee et al. 2014)
PAF26
Ac-rkkwfw (Lopez-Garcia et al. 2002)
PAS
GKPILFF (Woldetsadik et al. 2017)
pCLIP6
KVRVRVRVpP(pT)RVRERVK (Chen et al. 2017b)
pD-SP5
riPRPSPKMGV(pS)VS (Chen et al. 2017b)
PenetraMax
KWFKIQMQIRRWKNKR, L- and D- (Khafagy el et al. 2015)
Penetratin
RQIKIWFQNRRMKWKK (Derossi et al. 1994)
Pep-1
KETWWETWWTEWSQPKKKRKV cysteamide (Morris et al. 1997)
pepM
KLFMALVAFLRFLTIPPTAGILKRWGTI (Freire et al. 2014)
pepR
LKRWGTIKKSKAINVLRGFRKEIGRMLNILNRRRR (Freire et al. 2014)
Pept1
PLILLRLLRGQF (Marks et al. 2011)
Peptide 599
GLFEAIEGFIENGWEGMIDGWYGGGGRRRRRRRRRK (Alexander-Bryant et al. 2015)
Pep42
Cyclic CTVALPGGYVRVC (Kim et al. 2006)
PepNeg
SGTQEEY (Neves-Coelho et al. 2017)
PepFect6
Stearyl-AGYLLGK(εTMQ)INLKALAALAKKIL, PF6 (El-Andaloussi et al. 2011)
PepFect14
Stearyl- AGYLLGKLLOOLAAAALOOLL (Ezzat et al. 2011)
PG1
RGGRLCYCRRRFCVCVGR (Liu et al. 2013b)
pHLIP
AEQNPIY-WARYADWLFTTPLLLLDLALLV-DADEGT (Andreev et al. 2010)
PHPs
H6-H10 peptides (Kimura et al. 2017)
PIP1
RXRRXRRXRIKILFQNRRMKWKK (Ivanova et al. 2008)
Pip5e
RXRRBRRXRILFQYRXRBRXRB (Betts et al. 2012)
Pip6a
Ac-RXRRBRRXRYQFLIRXRBRXRB (Lehto et al. 2014)
POD
CGGG(ARKKAAKA)4 (Dasari et al. 2017)
PR9
FFLIPKG-R9 (Liu et al. 2013a)
PTD
YARVRRRGPRRR (Dong et al. 2016)
PTD3
R9-ETWWETWWTEW (Kizaka-Kondoh et al. 2009)
PTD4
YARAAARQARA (McCusker et al. 2007)
Poly-Arg
Most popular R7 – R12 (Mitchell et al. 2000, Futaki, 2006)
pVEC
LLIILRRRIRKQAHAHSK (Elmquist et al. 2001)
Pyrrhocoricin
VDKGSYLPRPTPPRPIYNRN (Otvos et al. 2000)
R4K1
Stapled Ac-RRRRKS*LHRS*LQDS (Speltz et al. 2018)
R6dGR
R6dGR (Wang et al. 2017)
R8
R8 (Wender et al. 2001)
R8-dGR
R8dGR (Liu et al. 2016b)
R9-H4A2
Ac-YR9-HAHAHH (Okitsu et al. 2017)
R6W3
R6W3 (Bechara et al. 2013)
R10W6
R10W6 (Bechara et al. 2013)
RA9
RRAARRARR (Alhakamy et al. 2013)
RALA
WEARLARALARALARHLARALARALRACEA (McCarthy et al. 2014)
RDP
CKSVRTWNEI IPSKGCLRVG GRCHPHVNGG GRRRRRRRRC (Xiao et al. 2017)
REDV
REDV (Yang et al. 2016)
RF
GLKKLARLFHKLLKLGC (Yang et al. 2014)
cRGDfC
Cyclic RGDfC (Wada et al. 2017)
iRGD
Cyclic CRGDKGPDC (Peng and Kopecek, 2015)
RGE
RGERPPR (Yu et al. 2017)
RH9
RRHHRRHRR (Alhakamy et al. 2013)
RL9
RRLLRRLRR (Alhakamy et al. 2013)
RL16
RRLRRLLRRLLRRLRR (Joanne et al. 2009)
RT53
RQIKIWFQNRRMKWKKAKLNAEKLKDFKIRLQYFARGLQV YIRQLRLALQGKT (Jagot-Lacoussiere et al. 2016)
RTP004
RKKRRQRRRG-K15-GRKKRRQRRR) (Lee et al. 2015a)
RV24
RRRRRRRRRGPGVTWTPQAWFQWV (Lo and Wang, 2012)
RVG
YTIWMPENPRPGTPCDIFTNSRGKRASNG (Kumar et al. 2007)
RVG-9R
YTIWMPENPRPGTPCDIFTNSRGKRASNGGGGRRRRRRRRR (Rassu et al. 2017)
RVG29
YTIWMPENPRPGTPCDIFTNSRGKRASNGGGGRRRRRRRRR (Villa-Cedillo et al. 2017)
RW9
RRWWRRWRR (Alhakamy et al. 2013)
RW16
RRWRRWWRRWWRRWRR (Jobin et al. 2013)
(RXR)4
(R-Ahx-R)4 (Saleh et al. 2010)
(rXr)4
(r-Ahx-r)4 (Vij et al. 2016b)
S155
VKKKKIKREI-KIAAQRYGRELRRMADEFHV (Haidar et al. 2017)
S4(13)-PV
ALWKTLLKKVLKAPKKKRKV (Mano et al. 2007)
SAP
VRLPPPVRLPPPVRLPPP (Pujals et al. 2006)
SAP(E)
VELPPPVELPPPVELPPP (Martin et al. 2011)
all-D-SAP
(vrlppp)3 (Pujals et al. 2007)
SAPSp-lipo
stearyl-GGGGHGAHEHAGHEHAAGEHHAHE (Suzuki et al. 2017)
SAR6EW
SAR6EW (Im et al. 2017)
sC18
GLRKRLRKFRNKIKEK (Oren et al. 1999)
(sC18)2
(GLRKRLRKFRNKIKEK)2 (Gronewold et al. 2017)
SMTP motif,
LRLLR (Fuselier and Wimley, 2017)
SPACE
Cyclic ACTGSTQHQCG (Hsu and Mitragotri, 2011)
SRCRP2-11
GRVEVLYRGSW (Tuttolomondo et al. 2017)
STR-KV
H3K3V6 (Pan et al. 2016)
SS-02
Dmt-r-FK (Alta et al. 2017)
SS-20
F-r-FK (Alta et al. 2017)
SS-31
r-Dmt-KF (Zhao et al. 2005)
SynB1
RGGRLSYSRRRFSTSTGR (Rousselle et al. 2000)
T2
LVGVFH (Kumar et al. 2012)
Tat(49-57)
RKKRRQRRR (Vives et al. 1997a)
Tat(48-60)
GRKKRRQRRRPPQ (Vives et al. 1997b)
Tat(44-57)
CGISYGRKKRRQRRR (Niesner et al. 2002)
Tat(37-72)
CFITKALGISYGRKKRRQRRRPPQGSQT-HQVSLSKQ (Fawell et al. 1994)
Tat analog
GRKKRRQR (Nguyen et al. 2008)
Tat-LK15
Tat-KLLKLLLKLLLKLLK (Peng et al. 2017a)
TCTP
MIIFRALISHKK (Bae et al. 2016)
TD-1
ACSSSPSKHCG (Chen et al. 2006)
TD2.2
SYWYRIVLSRTGRNGRLRVGRERPVLGESP (Heffernan et al. 2012)
TH peptide
GYLLGHINLHHLAHL-Aib-HHIL (Chen et al. 2017a)
TM2
PKKGSKKAVTKAQKKDGA (Kochurani et al. 2015)
Transportan
GWTLNSAGYLLGKINLKALAALAKKIL, TP (Pooga et al. 1998)
TP10
AGYLLGKINLKALAALAKKIL (Soomets et al. 2000)
TPk
VRRFkWWWkFLRR (Bahnsen et al. 2015)
Tpl
KWCFRVCYRGICYRRCRGK (Jain et al. 2015)
TPP
TKDNNLLGRFELSG (Gehrmann et al. 2014)
TT1
CKRGARSTA (Paasonen et al. 2016)
vAMP 059
INWKKWWQVFYTVV (Dias et al. 2017)
vCPP 0769
RRLTLRQLLGLGSRRRRRSR (Dias et al. 2017)
vCPP 2319
WRRRYRRWRRRRRWRRRPRR (Dias et al. 2017)
VDAC(1-26)
MAVPPTYADLGKSARDVFTKGYGFGL (Smilansky et al. 2015)
VP22
NAATATRGRSAASRPTQRPRAPARSASRPRRPVQ (Elliott and O’Hare, 1997)
V peptide
TVDNPASTTNKDKLFAVRK (Manosroi et al. 2014)
VT5
DPKGDPKGVTVTVTVTVTGKGDPKPD (Oehlke et al. 1997)
W(RW)4
W(RW)4 (Nasrolahi Shirazi et al. 2013)
Xentry
LCLR (Montrose et al. 2014)
X-pep
MAARLC (Adachi et al. 2017)
YKA
YKALRISRKLAK (Desai et al. 2014)
YTA2
YTAIAWVKAFIRKLRK (Lindgren et al. 2006)
YTA4
IAWVKAFIRKLRKGPLG (Lindgren et al. 2006)
Z2
FWIGGFIKKLKRSKLA (Chen et al. 2017c)
Z3
FKIKKFIGGLWRSKLA (Chen et al. 2017c)
Z12
KRYKNRVASRKCRAKFKQLLQHYREVAAAKSSENDRLRLLLK (Derouazi et al. 2015)
ZXR-1
FKIGGFIKKLWRSKLA (Chen et al. 2017c)
[/showhide]
Manage Consent
To provide the best experiences, we use technologies like cookies to store and/or access device information. Consenting to these technologies will allow us to process data such as browsing behavior or unique IDs on this site. Not consenting or withdrawing consent, may adversely affect certain features and functions.
Functional
Always active
The technical storage or access is strictly necessary for the legitimate purpose of enabling the use of a specific service explicitly requested by the subscriber or user, or for the sole purpose of carrying out the transmission of a communication over an electronic communications network.
Preferences
The technical storage or access is necessary for the legitimate purpose of storing preferences that are not requested by the subscriber or user.
Statistics
The technical storage or access that is used exclusively for statistical purposes.The technical storage or access that is used exclusively for anonymous statistical purposes. Without a subpoena, voluntary compliance on the part of your Internet Service Provider, or additional records from a third party, information stored or retrieved for this purpose alone cannot usually be used to identify you.
Marketing
The technical storage or access is required to create user profiles to send advertising, or to track the user on a website or across several websites for similar marketing purposes.